














Chapitres
Notice
Role du trafic membranaire dans la morphogenèse neuronale
- document 1 document 2 document 3
- niveau 1 niveau 2 niveau 3
Descriptif
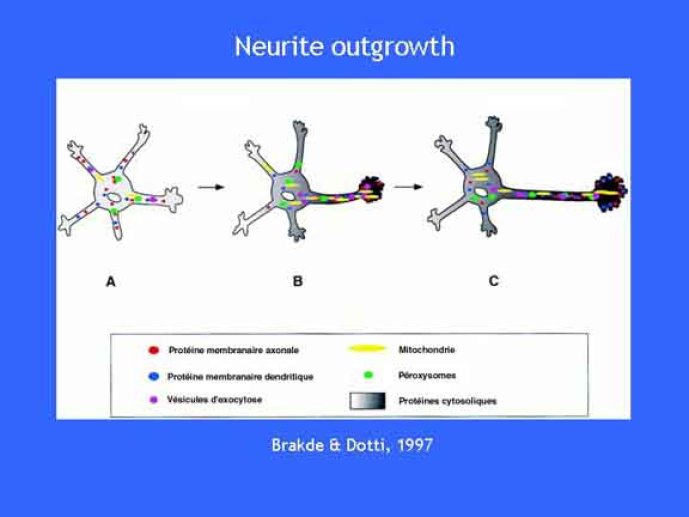
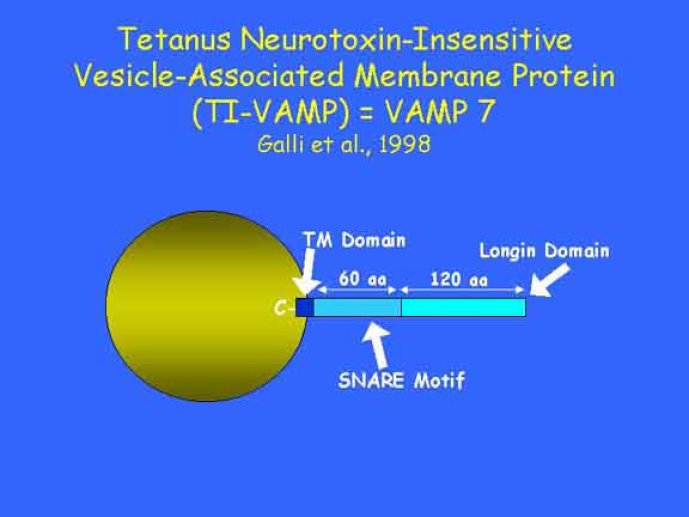
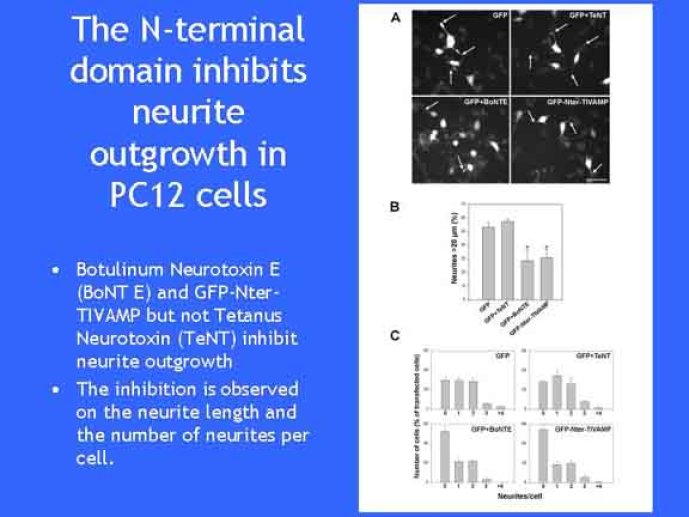
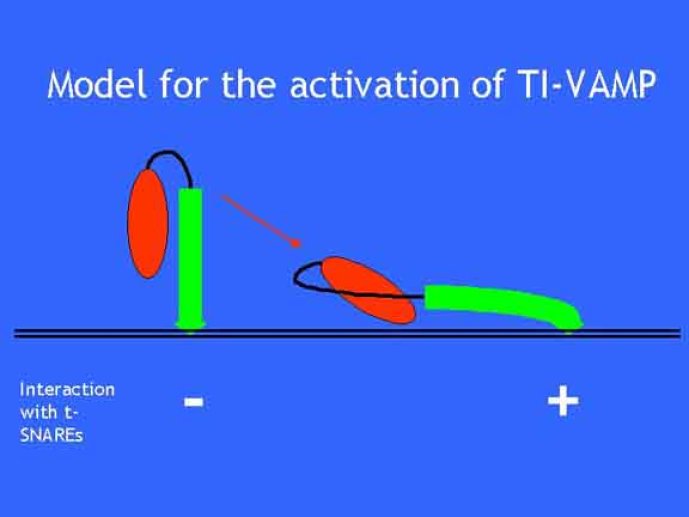
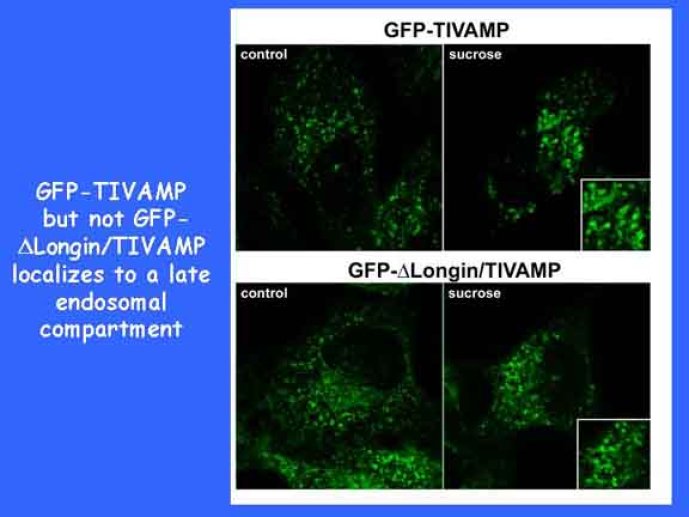
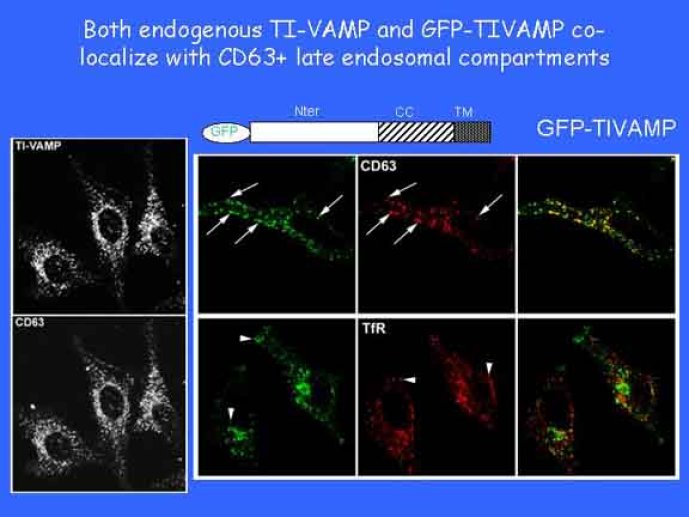
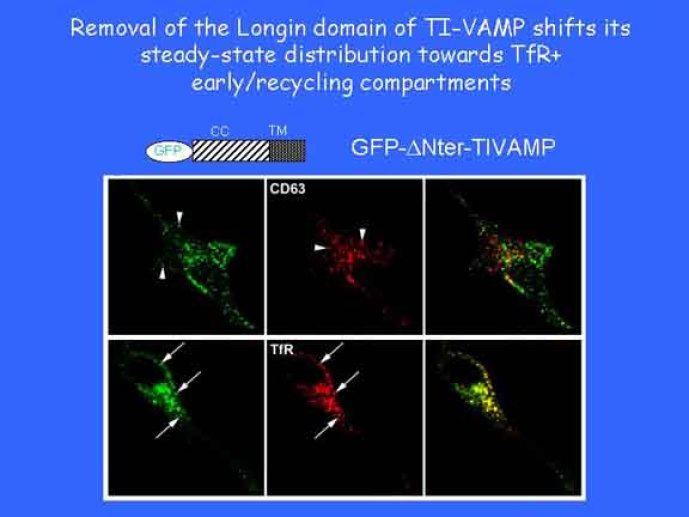
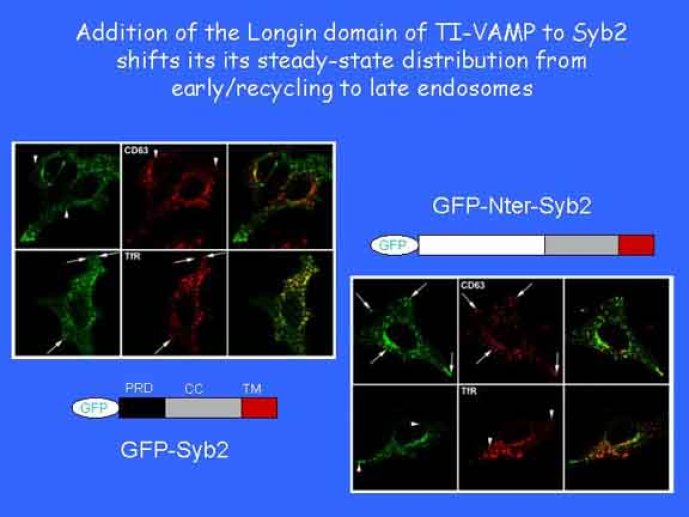
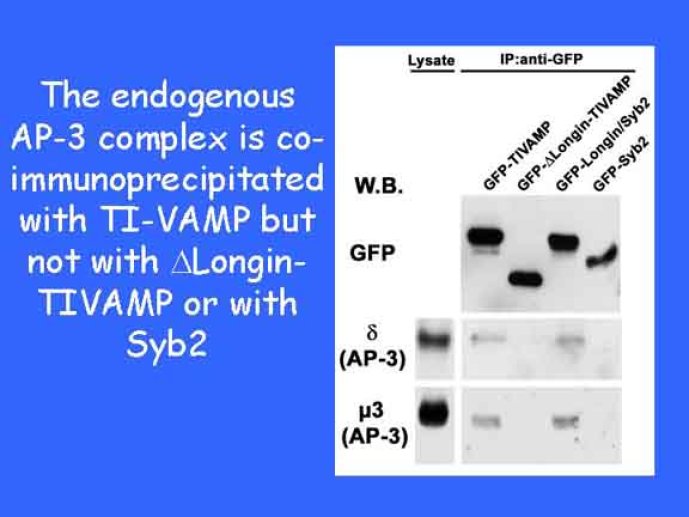
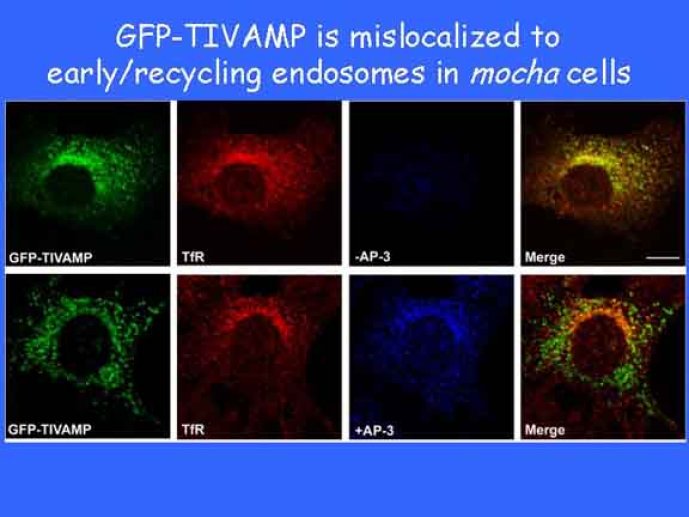
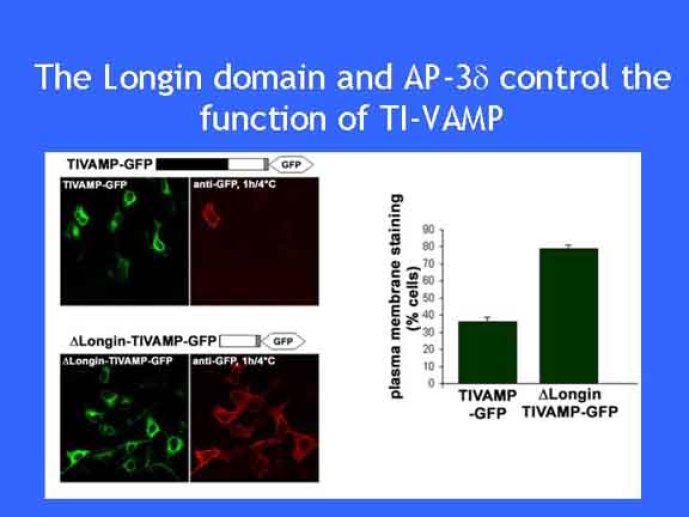
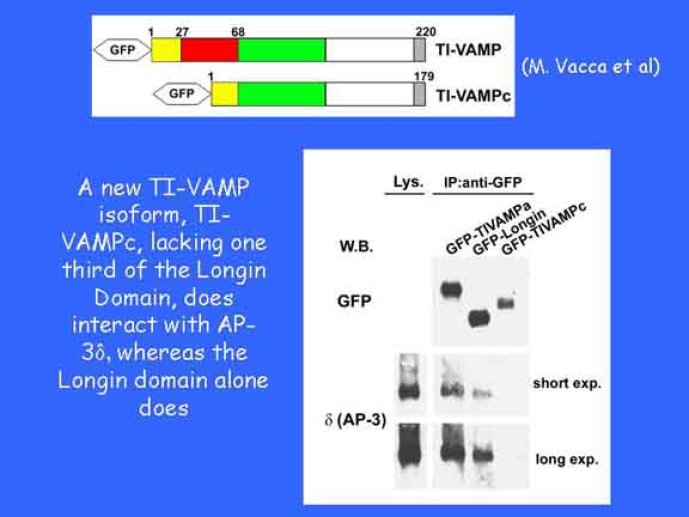
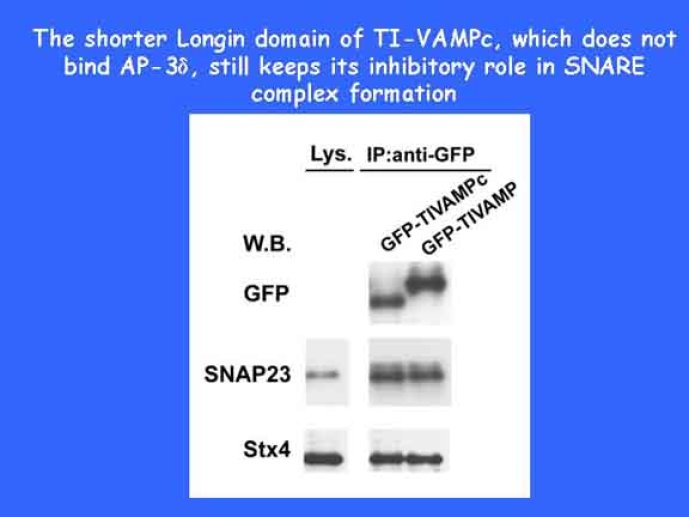
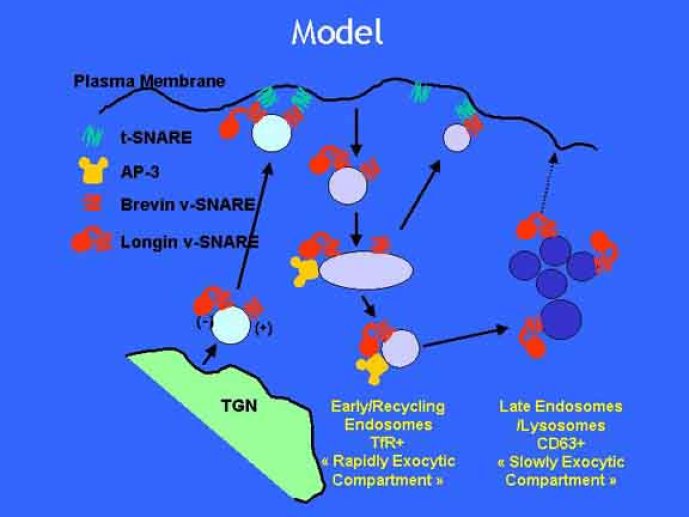
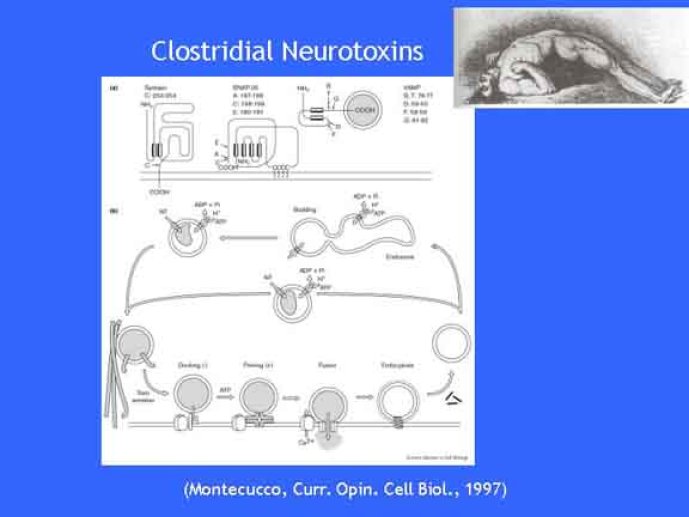
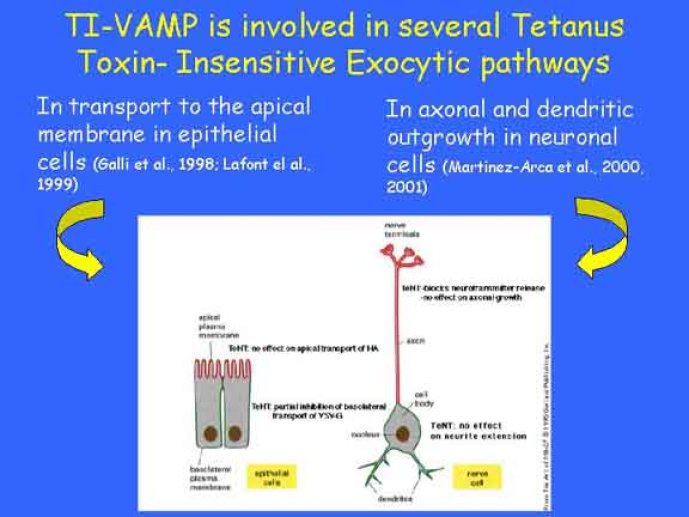
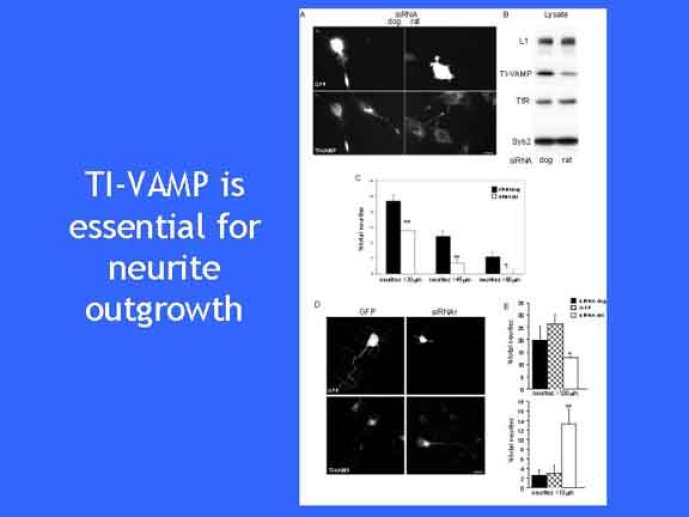
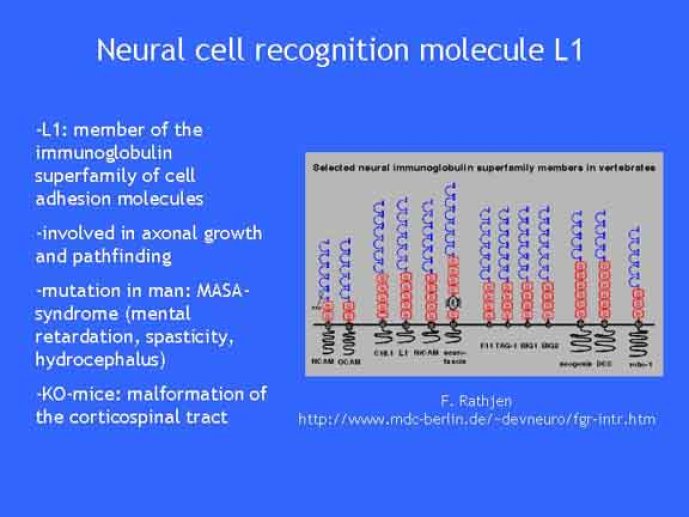
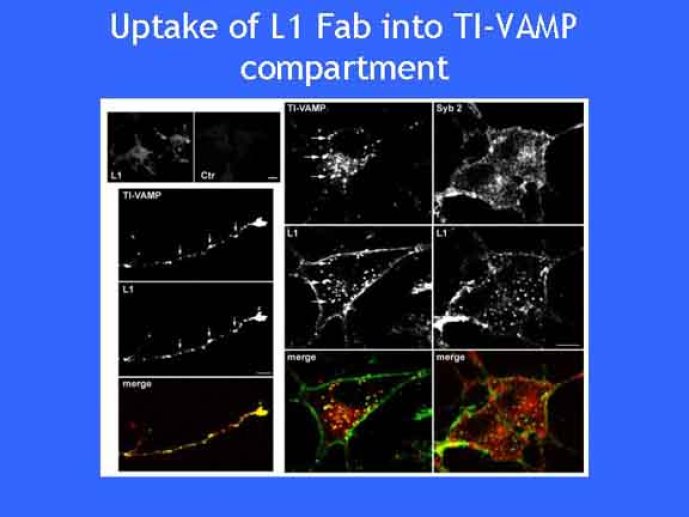
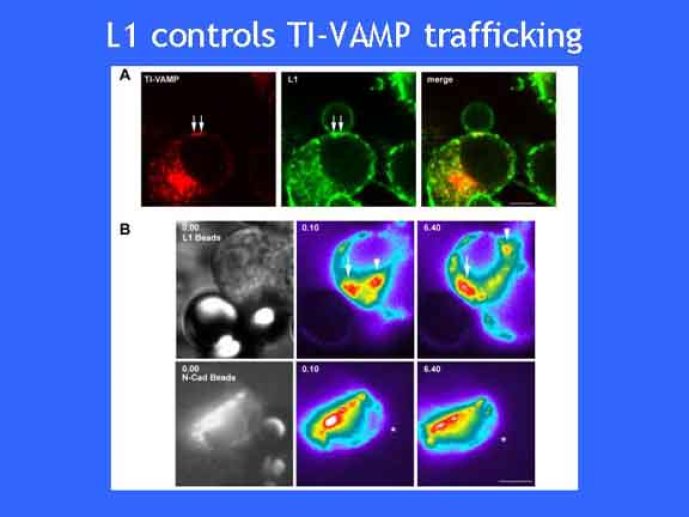
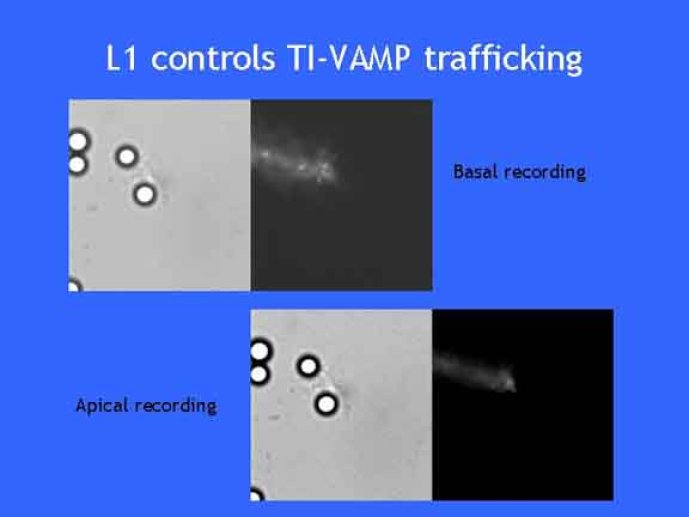
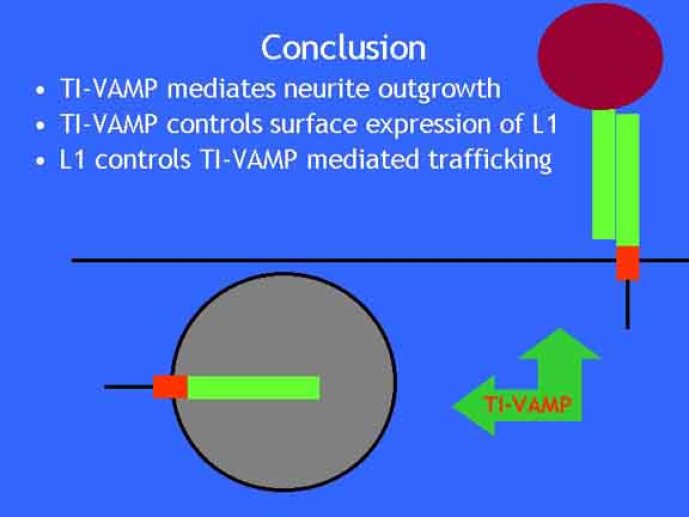
Le trafic membranaire est à la base des processus sécrétoires des cellules neuronales et non-neuronales mais son rôle dans l'établissement dans la différenciation neuronale est encore mal établi. Notre objectif est de comprendre comment le trafic membranaire, plus particulièrement l'exocytose et l'endocytose, participent à la morphogenèse neuronale. L'importance des protéines SNAREs dans l'exocytose et dans chaque étape du trafic membranaire est maintenant bien établie. Dans les neurones, la voie d'exocytose des vésicules synaptiques, responsable de la libération des neurotransmetteurs, met en jeu la protéine vésiculaire synaptobréviné 2 (ou VAMP 2, un v-SNARE) qui forme un complexe avec ses SNAREs cibles à la membrane plasmique : SNAP25 et la syntaxine 1, qui, ensemble, forment le t-SNARE). La formation du complexe v-/t-SNARE permet la fusion des bicouches lipidiques de la membrane vésiculaire et de la membrane plasmique 1. Dans l'équipe, nous avons mis en évidence Tetanus neurotoxin Insensitive-VAMP (TI-VAMP) un nouveau membre de la famille VAMP/brévine. Contrairement aux VAMPs 1, 2, et 3 qui sont clivées par la toxine tétanique et les neurotoxines botuliques B, D, F et G, TI-VAMP est insensible aux neurotoxines 2. Nous avons montré précédemment que TI-VAMP se concentre à l'extrémité des cônes de croissance de l'axone et des dendrites 3 et est impliquée dans la croissance neuritique dans les cellules PC12 et les neurones. En effet, l'expression de l'extrémité amino-terminale de TI-VAMP appelé domaine Longin 4 inhibe la croissance neuritique alors que l'expression d'une forme délétée du domaine Longin l'active 5,6. Nous montrons que le domaine Longin contrôle la capacité de TI-VAMP de former des complexes SNAREs avec SNAP25 et syntaxin1 et régule la localisation de TI-VAMP en intéragissant avec l'adaptateur de la clathrine AP-3. En conséquence, ce domaine contrôle simultanément l'activité de TI-VAMP et son ciblage. De plus, l'extinction de l'expression de TI-VAMP par interférence d'ARN bloque la croissance neuritique dans les cellules PC12 et les neurones. Nos résultats récents montrent que TI-VAMP est impliquée dans le trafic de la protéine d'adhésion cellulaire L1 et dans l'adhésion dépendante de L1. L1 est un membre de la superfamille des protéines à domaines immunoglobulines qui a été impliqué dans le développement du cerveau et dont des mutations entraînent des malformations cérébrales 7. Par ailleurs, nous montrons que l'adhésion dépendante de L1 contrôle le trafic membranaire dépendant de TI-VAMP, établissant ainsi une convergence entre trafic membranaire et adhésion cellulaire. L'ensemble de ces résultats démontre que la voie de trafic membranaire dépendante du v-SNARE TI-VAMP joue un rôle fondamental dans la morphogenèse neuronale.
Thème
Documentation
Liens
Dans la même collection
-
Jean-Antoine Girault: Les jonctions paranodales
GiraultJean-AntoineDans le cadre de la journée organisée par André Calas en hommage à Andrée Tixier-Vidal, Jean-Antoine Girault nous parle des interactions entre cellules gliales et neurones au niveau des noeuds de
-
Trafic intraneuronal des récepteurs couplés aux protéines G
Conférence donnée par Bertrand Bloch à l'occasion du jubilé en l'honneur de Andrée Tixier-Vidal.
-
Les récepteurs à dépendance : Interface entre apoptose et développement du système nerveux
MehlenPatrickLa recherche des mécanismes de réception de signaux extracellulaires par une cellule a jusqu'ici toujours été considérée selon le schéma classique suivant: un ligand fixe un récepteur, ce récepteur
-
Dynamique de l'organisation des récepteurs du glutamate
Dans le cadre du jubilé organisé en l'honneur de Andrée Tixier-Vidal, Daniel Choquet a donné une conférence sur la dynamique de l'organisation des récepteurs au glutamate dans la membrane neuronale.
-
Discours de Andrée Tixier-Vidal
Andrée Tixier-Vidal conclue la journée donnée en son honneur en retraçant les différentes étapes de sa brillante carrière.


Sur le même thème
-
Diplôme Universitaire - Apprendre à apprendre
Présentation de la formation
-
L'exploration mentale #4 – Intelligence émotionnelle et la compréhension de ses granularités
LacroixÉricPorlierChristopheL'exploration mentale #4 – Intelligence émotionnelle et compréhension de ses granularités
-
L'exploration mentale #3 – La visualisation dans la performance sportive
LacroixÉricPorlierChristopheL'exploration mentale #3 – La visualisation dans la performance sportive
-
Une minute avec Amélie Aussel
AusselAmélieAmélie Aussel est chercheuse au sein de l’équipe-projet Mnemosyne du centre Inria de l’université de Bordeaux.
-
The coldness of the stone and the lightness of the bird: a memorial garden for the victims of Novem…
AccarainMadeleineTalk by Madeleine ACCARAIN (Wagon Landscaping, Paris), as part of the workshop "Memory, Place, and Material Culture", organized by John SUTTON, 2022-2023 research fellow at the Paris IAS, Emeritus
-
mark edmonds, building and belonging: architecture and memory in neolithic orkney
EdmondsMark R.SuttonJohnTalk by Mark EDMONDS, (Archeologie, York), in the context of the workshop "Memory, Place, and Material Culture", organized by John SUTTON, 2022-2023 research fellow at the Paris IAS, Emeritus
-
La théorie des graphes pour comprendre la réorganisation cérébrale après un coma.
AchardSophieLa théorie des graphes pour comprendre la réorganisation cérébrale après un coma.
-
Topological insights in neuroscience
Hess BellwaldKathrynOver the past decade, and particularly over the past five years, research at the interface of topology and neuroscience has grown remarkably fast. Topology has, for example, been successfully applied
-
Thomas VERGINE (Unicaen, Craham), Salives (Côte-d’Or), Morphogenèse d’une agglomération d’origine c…
VergineThomasSalives est un site particulièrement représentatif de la morphogenèse d’une petite agglomération bourguignonne autour d’un site élitaire au cours du Moyen Âge. Le village actuel comporte un patrimoine
-
Consommation ou sobriété : comment notre cerveau nous permet de choisir ?
GranonSylvieAu cours de cet exposé nous verrons comment les connaissances actuelles sur le fonctionnement cérébral et la prise de décision permettent d’expliquer comment se font les choix, leur diversité et ce
-
The Iseult project : a brain spacetime odyssey
Over the past ten years, Bio-Imaging translational research has experienced an unprecedented expansion: ten years of challenges, collaborations, success stories and significant advances, thanks
-
brains on drugs: lessons from the 3rd reich
Norman OHLER (Berlin – author of "Blitzed, Drugs in the Third Reich"): "Brains on Drugs: Lessons from the Third Reich" "The Brains that pull the Triggers - 3rd Paris Conference on Syndrome E":